
Theory & Practice: The Extensor Paradox in Running
The original article (below) on the topic at hand, was published in Biomechanics of Distance Running in 1990. As you read the data and conclusions from this research you’ll see that the scientific community was not ready to accept the idea of the role of gravity as a leading force in running. A classical vision of gravity strictly as a vertical force was predominant in the scientists’ minds and didn’t allow them to look at the facts from a different perspective. The most important thing there, a relationship between extensor muscles and gravity as one non-conflicting system with reciprocal coordination between them, was overlooked.
The Data
The data in this article clearly demonstrates Nature’s wisdom of coexistence, when one force yields to the other to allow them both be used to their fullest. In the Pose Method the concept of gravity as a leading force in forward movement is the most fundamental one, and the data from the extensor’s paradox article below confirm this.
I would like to specifically point out for you the data showing when the quad muscles cease any electrical activity. According to the research data, it happens immediately after the mid-stance, when according to the traditional point of view the so-called “push off” efforts are supposed to be exerted.
This conflicting information brought the authors to this particular name of the article. The commonly accepted understanding of the leg extension as a forward propulsive force in running did not get any support by the data provided by this research. But, at the same time, with this data available, the researchers did not come to any conclusions that should have pointed out the role of gravity in running.
The most important thing there, a relationship between extensor muscles and gravity as one non-conflicting system with reciprocal coordination between them, was overlooked.
The Logic
In the absence of pre-existing standards and guidelines, we must step away from the microscope and look at the bigger picture. Figuring out the hierarchy of the existing forces and how they work/interact elsewhere and everywhere allows us to lay down the ground rules. This initial sorting of already known facts is essential in formulating a concept.
Currently our understanding of the force of gravity is limited and yet we know just enough to understand that it is the glue that holds everything together. Thus it is considered the leading force. If we accept it as such in relation to our entire planet, then we must accept gravity as the leading force in horizontal movement in running as well, all other forces are subordinate.
Jumping ahead to the work of muscles and our entire body framework of bones, connective tissues, etc it is logical to suggest that when we see a muscle group cease activity that it happens so specific muscle behavior does not interfere with the work of gravity but falls in line with it. It is easy to say that this logic has been established from the beginning of biological life on Earth. From this point of view, our conscious efforts to produce the forward propulsion were “ignored” by Nature.
The Practice
Some perception of muscle efforts on support, which we have during the support time right before and during mid-stance, gives an illusion of this “push off” happening. Most runners sincerely believe in ‘push off efficiency’ and its necessity in order to run, because of their perception and deceptive visual appearance. The fact is that we feel tension interpreted as a push simply because we arrive to the single point of support with our entire body weight on it for a fleeting moment in time.
Try this. Stand in the running pose and start falling forward. Now push off. Be honest with yourself instead of just trying to prove me wrong. Could you push off? No.
Our common sense is based on and is limited by our understanding of the subject, and hence is a very deceptive thing that often doesn’t coincide with abstract logic, which we have to use in order to see the hidden reality of functioning of systems. For this matter we have to use the system of reference of Nature, applying scientific terminology, according to which Gravity is a predominant force by all accounts. Then and only then we’ll be able to see how the forces interact within the hierarchically structured system, each with its own space and time of involvement in the action of running.
BIOMECHANICS OF DISTANCE RUNNING
Chapter 6. Muscle Activity in Running. The Extensor Paradox Experiment
by Irene S. McClay, Mark J. Lake, Peter R. Cavanagh
It is well known that knee flexion occurs just before and immediately after footstrike during running to cushion the impact of landing (Milliron & Cavanagh, this volume). Once the downward movement of the center of gravity associated with this cushioning phase has finished, knee extension begins and the propulsive phase of the cycle continues.
There is evidence from Brandell (1973) and Mann and Hagy (1980b) that the quadriceps are generally silent during the phase of knee extension following the cushioning. Few experiments have focused on this puzzling aspect of knee joint action during running. The purpose of the experiment described in this section was to examine the activity of the three heads of the quadriceps that are amenable to surface recording during distance running and to simultaneously measure the angle of the knee joint.
Subjects and Speed
Six male recreational runners, ages 19 to 26, experienced in treadmill running with no history of recent injury, volunteered for the study. Each subject ran at a constant speed of 4.0 m · S-¹ on a motorized treadmill. This speed was chosen as it was in the middle of the range used by previous workers.
Equipment and Method of Analysis
To investigate knee extensor muscle activity during the stance phase of running, EMG of the vastus medialis, vastus lateralis, and rectus femoris muscles of one leg were recorded using a battery-powered GCS 67 Electromyographic Processor. Silver-silver chloride electrodes with on-site preamplifiers were placed in the middle of the muscle belly after thorough preparation of the skin. An inertia switch attached to the heel was used to define the cycle endpoints and knee angle was recorded simultaneously with a self-aligning ULGN-67 Electrogoniometer. This design compensates for errors in placement and does not assume a fixed center of rotation for the joint. The electrogoniometer was calibrated for knee angle by comparing voltage output against knee angle measured by a protractor.
The EMG processor, together with the goniometer and footswitch signals, were interfaced with an SMS 1000 computer, which sampled at a rate of 500 Hz per channel. The raw EMG signal was prefiltered using a high pass filter of 75 Hz cut-off frequency. Custom software allowed for storage, processing, and display of the data. An example of the raw data for the complete 5-second sampling period is shown in Figure 6.6a, and the region surrounding footstrike is shown with greater resolution in Figure 6.6b.
")
Figure 6.6a. A 5-s raw experimental record.
")
Figure 6.6b. A portion of the same experimental record surrounding footstrike shown with greater time resolution.
Five-second samples were collected after each subject had undergone a warm-up period at the test speed. This allowed at least six full cycles of running to be recorded for each individual. For each period of stance, the phasic activity of all three muscles was subjectively determined by comparison with a noise-free baseline. Data from six footstrikes were examined, and mean values were obtained for the time at which rectus femoris, vastus lateralis, and vastus medialis muscle activity ceased. The beginning and end times of the knee extension phase following initial flexion were also determined.
Results
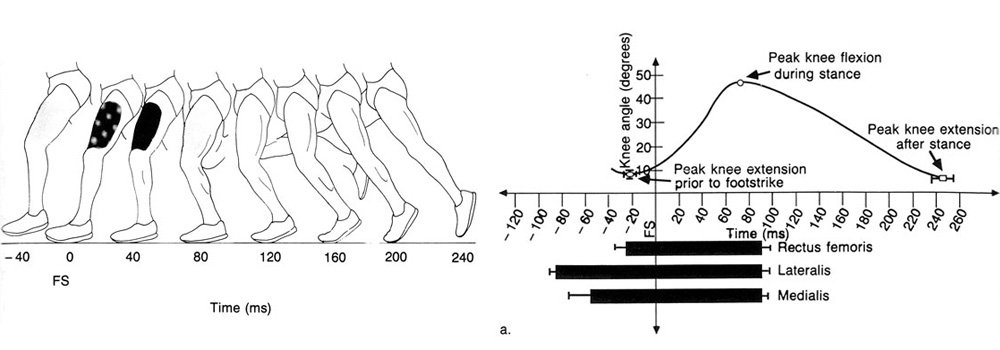
Figure 6.7a illustrates the mean results of six footstrikes for a typical subject. It can be seen that approximately 85 milliseconds before footstrike, muscle activity begins while knee extension is under way. Vastus lateralis is the first to show activity, some 25 milliseconds before vastus medialis and 60 milliseconds before rectus femoris. This period of muscle activity appears to help in stabilizing the leg in preparation for footstrike. All three muscles are active through footstrike while knee flexion occurs, but they cease activity simultaneously approximately 20 milliseconds after peak knee flexion has been achieved. In this subject knee extension continues for a further 150 milliseconds.

Figure 6.7a. Results of phasic quadriceps EMG and knee angle for a typical subject averaged over six footstrikes.

Figure 6.7b Ensemble average results of six subjects of the relationship between phasic quadriceps EMG and knee angle. The values of peak knee extension prior to footstrike, peak knee flexion during stance, and peak knee extension after stance have been joined by straight lines as the mean curve was not determined.
The mean results for the group as a whole are presented in Table 6.1 and shown schematically in Figure 6.7b. The mean time of knee extension that was not accompanied by quadriceps EMG was 133.7 milliseconds (SD = 16.5).
| Flexion extension duration | Mean all muscle off after peak flexion | Mean duration of silence during extension | |
| Mean for group ±SD | 162.8 19.5 | 29.2 10.4 | 133.7 16.5 |
These results are further illustrated in Figure 6.8, where electrical activity is indicated by the presence of shading over the muscle. The amplitude of the activity is also schematically indicated by the intensity of the shading. The large amount of knee extension that occurs in the absence of muscle activity is readily apparent from this figure.

Figure 6.8. The amplitude of EMG activity throughout the stance phase of running. (The intensity of shading indicates relative amount of activity.)
Discussion
For the group of runners examined in this study, it is clear that the quadriceps cease their activity shortly after peak stance phase knee flexion has occurred. A phase of knee extension of approximately 130 milliseconds continues without the assistance of the quadriceps. The function of the quadriceps must therefore be described as principally controlling the descent of the body center of gravity after landing. Certainly they help to initiate knee extension, but they rapidly become quiescent when knee extension has been under way for only about 30 milliseconds, a time during which less than 5 degrees of extension has been achieved. The duration of electrical silence in extension is large enough to exclude the possibility that electromechanical delay (EMD) between EMG activity and force production may explain the paradox. EMD time in concentric muscle action has been determined to be 40 to 55 milliseconds (Cavanagh & Komi, 1979; Norman & Komi, 1979), and in rapid movements it may be possible for EMG activity to have terminated before force can be detected.
A reasonable hypothesis may be that hip extensor action during the second half of the stance phase is causing the knee joint to extend. However, if one examines the co-activation of the quadriceps and hamstrings in Figure 6.3, it is apparent that many investigators have found these muscle groups to cease activity at about the same time in the cycle. Neither does there appear to be a prolonged period of gluteus maximus activity that would provide an explanation. Figure 6.4 indicates that the last extensor muscle to cease activity during stance appears to be the gastrocnemius, which is of course also a knee flexor. Because only the quadriceps were measured in the present study, it is not possible to say with certainty what patterns of activity were exhibited in other muscles in these particular subjects. These experiments have, however, shown that the notion of an extensor thrust-with plantar flexors, knee extensors, and hip extensors all being active in late support to generate forward and upward thrust – is in need of modification. They also indicate that the problem is worthy of further investigation using a kinetic approach in addition to multi-channel EMG so that the joint moments can be determined.
References
- Basmajian, J.V., & Deluca, C.J. (1985). Muscles alive (5th ed.). Baltimore: Williams and Wilkins.
- Brandell, B.R. (1973). An analysis of muscle coordination in walking and running gaits. In S. Cerquiglini, A. Venerando, & J. Wartenweiler (Eds.), Medicine and Sport: Biomechanics III (pp. 278-287). Basel, Switzerland: Karger.
- Carlet, M. (1872). Essai experimental sur la locomotion humaine: Etude de la marche [Experimental test on human locomotion: Study of walking]. Annales des Sciences Naturelles, Sect. Zool., XV.
- Cavagna, G.A. (1977). Storage and utilization of elastic energy in skeletal muscle. Exercise and Sport Sciences Reviews, 5, 89-129.
- Cavanagh, P.R., & Komi, P.V. (1979). Electromechanical delay in human skeletal muscle under concentric and eccentric contractions. European Journal of Applied Physiology, 42, 159-163.
- Cohen, H.L., & Brumlik, J. (1968). A manual of electroneuromyography. New York: Harper and Row.
- Elliot, B.C., & Blanksby, B.A. (1979). The synchronization of muscle activity and body segment movements during a running cycle. Medicine and Science in Sports, 11(4), 322-327.
- Grieve, D.W., Pheasant, S., & Cavanagh, P.R. (1978). Prediction of gastrocnemius length from knee and ankle joint posture. In E. Asmussen & K. Jorgensen (Eds.), Biomechanics VI-A (pp.405-412). Baltimore: University Park.
- Hubbard, A.W. (1939). An experimental analysis of running and of certain differences between trained and untrained runners. Research Quarterly of the American Association of Health and Physical Education, 10(3), 28-38.
- Hudgkins, C.V., & Stetson, R.H. (1932, July 15). A unit for kymographic recording. Science, p. 60.
- Kramer, H., Kuchler, G., & Brauer, D. (1972). Investigations of the potential distribution of activated skeletal muscles in man by means of surface electrodes. Electromyography and Clinical Neurophysiology, 12, 19-26.
- MacIntyre, D.L., & Robertson, D.G.E. (1987). EMG profiles of the knee muscles during treadmill running. In Bengt Jonsson (Ed.), Biomechanics X-A (pp.289-294). Champaign, IL: Human Kinetics.
- Mann, R.A., & Hagy, J.L. (1980a). Biomechanics of walking, running, and sprinting. American Journal of Sports Medicine, 8(5), 345-350.
- Mann, R.A., & Hagy, J.L. (1980b). Running, jogging and walking: A comparative electromyographic and biomechanical study. In J.E. Bateman & A. Trott (Eds.), The foot and ankle (pp.167-175). New York: Thieme-Stratton.
- Marey, E.J. (1972). Movement. New York: Arno. (Original work published 1895)
- Nilsson, J., Thorstensson, A., & Halbertsma, J. (1985). Changes in leg movements and muscle activity with speed of locomotion and mode of progression in humans. Acta Physiologica Scandinavica, 123, 457-475.
- Norman, R.W., & Komi, P.V. (1979). Electromechanical delay in skeletal muscle under normal movement conditions. Acta Physiologica Scan dinavica, 106, 241-248.
- Norman, R.W., Nelson, R.C., & Cavanagh, P.R (1978). Minimum sampling time required to extract stable information from digitized EMGs. In E. Asmussen & K. Jorgensen (Eds.), Biomechanics VI-A (pp.237-243). Baltimore: University Park.
- Pare, E.B., Stern, J.T., & Schwartz, J.M. (1981). Functional differentiation within the tensor fasciae latae. Journal of Bone and Joint Surgery, 63-A(9), 1457-1471.
- Schwab, G.H., Moynes, D.R. Jobe, F.W., & Perry, J. (1983). Lower extremity electromyographic analysis of running gait. Clinical Orthopedics and Related Research, 176, 166-170.
- Warfel, J.H. (1974). The extremities (4th ed.). Philadelphia: Lea & Febiger.
- Winter, D.A. (1979). Biomechanics of human movement. New York: John Wiley & Sons.
- Zuniga, E.M., Truong, X.T., & Simons, D.G. (1969). Effects of skin electrode position on averaged electromyographic potentials. Archives of Physical Medicine and Rehabilitation, 50, 264-271.
About the Author

Dr. Nicholas Romanov is the developer of the Pose Method®. A passionate proponent of higher level of education in athletics, Dr. Romanov dedicated his entire career to sports education, scientific research and coaching. An Olympic Coach and a bestselling author, Dr. Romanov has taught on all continents and visited almost every country in the world.
[ Click here to learn more ]
CONTINUING EDUCATION FOR HEALTH + FITNESS PROFESSIONALS
Pose Method® of Running: A Master Course on Running is approved for 20 contact hours towards continuing education for Certified CrossFit Trainers, Board Certified Athletic Trainers and Physical Therapists.

Leave a Reply
Want to join the discussion?Feel free to contribute!